| Akiko IWANAGA's web page |
真正粘菌の細胞質伝達の直線順位の進化
ユニークな粘菌の性
岩波生物学事典によると、オスとは「雌雄異体の生物種において精巣をもつ動物個体,植物(または分化の程度の低い動物)では小配偶子をつくる個体」、メスは「卵を生ずる個体,または(下等な分化の低い種類では)大配偶子を形成する個体.」と定義されてる。
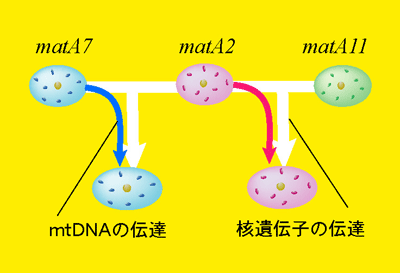 性の定義は様々あるが、本研究では雌雄の定義を次世代への細胞質遺伝子伝達に寄与するもの(メス)としないもの(オス)として考えた。例えば、多くの動物ではミトコンドリアは母方から受け継がれ、父方のものは受け継がれない。真正粘菌Physarum Polycephalumも配偶子が接合するときにミトコンドリアが片親遺伝する。ただし、真正粘菌は非常に多くの性(接合型)を持っているという点で他の2性の生物と異なっている。
性の定義は様々あるが、本研究では雌雄の定義を次世代への細胞質遺伝子伝達に寄与するもの(メス)としないもの(オス)として考えた。例えば、多くの動物ではミトコンドリアは母方から受け継がれ、父方のものは受け継がれない。真正粘菌Physarum Polycephalumも配偶子が接合するときにミトコンドリアが片親遺伝する。ただし、真正粘菌は非常に多くの性(接合型)を持っているという点で他の2性の生物と異なっている。では、3つ以上の性が存在する場合、ミトコンドリアはどのように伝達されるのだろうか。 実は、接合する相手によってミトコンドリアを伝達するか否かが変わってくる。
例えば、x,y,zという3つの接合型がある場合、 xとyが接合するときにはxからミトコンドリアが伝達される(xがメス、yがオス)。 が、yとzが接合するときにはyからミトコンドリアが伝達される(yがメス、zがオス)。
つまり、yは接合相手によってオスになったりメスになったりすると言うことできる。このミトコンドリアの伝達の優位性はphysarumの核に存在するのmatAという遺伝子が支配しており、15個の複対立遺伝子が存在することがわかっている。伝達の優位性は、
であることが交配実験によって確かめられた。matA7の核遺伝子を配偶子のミトコンドリアは必ず受け継がれ、matA6のミトコンドリアは常に排除される*。このようなミトコンドリアの伝達の直線的な順位関係はどのようにして、またどのような条件によって成立するのだろうか?
*接合時に双方のミトコンドリアが融合する現象が知られている。必ずしも劣位のミトコンドリアが全て排除されるとは限らない。
2つの遺伝子座
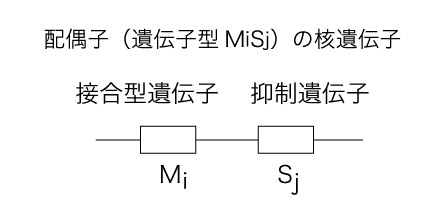 モデル作成にあたって、 接合型遺伝子M(配偶子の接合型(性)を決定する遺伝子)と、抑制遺伝子S(接合の際に相手のミトコンドリアDNA伝達を抑制する遺伝子)がそれぞれの配偶子の核遺伝子にコードされていると仮定した。それぞれの対立遺伝子をMi,Sjと呼ぶことにする。どちらの遺伝子もn個の対立遺伝子を持ち、2つの配偶子の接合型遺伝子座が異なっていればそれらは接合する。さらにそれぞれの配偶子が持つ抑制遺伝子座によって次世代にどちらのミトコンドリアが伝達されるかが決定される。
モデル作成にあたって、 接合型遺伝子M(配偶子の接合型(性)を決定する遺伝子)と、抑制遺伝子S(接合の際に相手のミトコンドリアDNA伝達を抑制する遺伝子)がそれぞれの配偶子の核遺伝子にコードされていると仮定した。それぞれの対立遺伝子をMi,Sjと呼ぶことにする。どちらの遺伝子もn個の対立遺伝子を持ち、2つの配偶子の接合型遺伝子座が異なっていればそれらは接合する。さらにそれぞれの配偶子が持つ抑制遺伝子座によって次世代にどちらのミトコンドリアが伝達されるかが決定される。Suppression Power model
このモデルでは、抑制遺伝子はその対立遺伝子ごとに接合相手のミトコンドリア伝達を抑制する力が異なっていることを仮定した。抑制遺伝子座S1が最も強い抑制力を持ち、Sn が最も弱い抑制力である(S1>S2>...>Sn)。
 これらの抑制遺伝子を持つ配偶子同士の接合と接合子へのミトコンドリアDNA(mtDNA) について考えよう。
まず、出会った配偶子のうち、お互いの接合型が異なれば接合をする。
接合可能な場合、2つの配偶子の持つ抑制遺伝子が異なれば抑制力の強い配偶子が接合相手の細胞質の伝達を阻止し、抑制力が同じであれば(同じ抑制遺伝子を持っていれば)、両方から細胞質が伝達される(heteroplasmy)。
この時、細胞質間のコンフリクトが起こり、接合子の適応度はコンフリクトを起こしていないものよりも下がる(heteroplasmic dippression)。細胞質コンフリクトという現象が接合子の適応度にどの程度の影響を及ぼすか詳細はわかっていないが、性の進化の研究で使用されている仮定である(Hurst and Hamilton,1992)。
これらの抑制遺伝子を持つ配偶子同士の接合と接合子へのミトコンドリアDNA(mtDNA) について考えよう。
まず、出会った配偶子のうち、お互いの接合型が異なれば接合をする。
接合可能な場合、2つの配偶子の持つ抑制遺伝子が異なれば抑制力の強い配偶子が接合相手の細胞質の伝達を阻止し、抑制力が同じであれば(同じ抑制遺伝子を持っていれば)、両方から細胞質が伝達される(heteroplasmy)。
この時、細胞質間のコンフリクトが起こり、接合子の適応度はコンフリクトを起こしていないものよりも下がる(heteroplasmic dippression)。細胞質コンフリクトという現象が接合子の適応度にどの程度の影響を及ぼすか詳細はわかっていないが、性の進化の研究で使用されている仮定である(Hurst and Hamilton,1992)。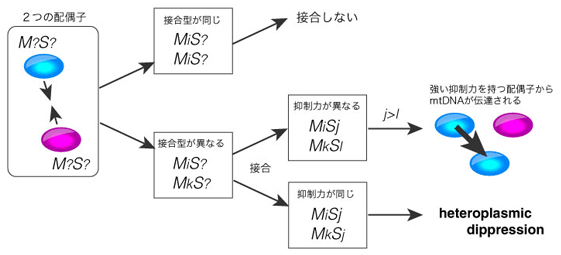
Suppression Power Model からわかること
2つの遺伝子の対立遺伝子の数をそれぞれ3つと仮定すると、集団中には計9種類の遺伝子型が存在することになる。では、ミトコンドリア遺伝子の伝達は接合型別に見るとどのようになっているだろうか。 この時点では、それぞれの接合型の中に3種類の抑制遺伝子が存在するので、接合型別のmtDNA伝達方向は決定できない。この状態からシミュレーションを行い、集団中の各遺伝子型の配偶子頻度の変化を調べたところこのような結果になった。
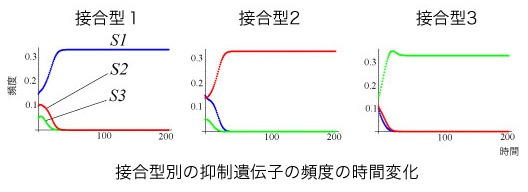
一接合型1は抑制遺伝子1で、接合型2は抑制遺伝子2で、接合型3は抑制遺伝子3で占められている。先程の初期状態と比較すると最終状態では各接合型一種類の抑制遺伝子で占められることがわかる。この3つの遺伝子型以外のものは消滅している。シミュレーションを多数回行った結果、各接合型の中の抑制遺伝子頻度分布がどのようであっても最終的に1つの接合型を単一の抑制遺伝子が占める状態に収束し、1つの接合型に複数の抑止遺伝子が混在することはなかった。
接合型別に見ると、
という線形の順位関係が成立します。
しかし、接合型遺伝子と抑制遺伝子との間に組み替えがある場合は結果が異なる。 組み換え率rが大きい時は9種類全ての遺伝子型が集団中に残る。この場合は接合型別の細胞質遺伝子の抑制方向が決定されず、直線順位は表れない。線形順位が成立する条件はr<alpha/n^2(alpha:heteroplasmic dippression, n:接合型の数)というシンプルな関係式で表すことができる。この式から、接合型遺伝子座と細胞質伝達抑制遺伝子座がタイトにリンクしている場合に線形順位が成立することがわかる。
Site-Specific Nuclease Model
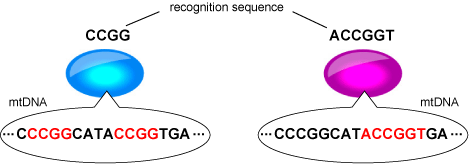 このモデルでは、認識配列に従ったDNAの切断による接合相手由来の細胞質遺伝子伝達抑制を仮定した。
ヌクレアーゼによって接合相手のミトコンドリアDNA(mtDNA)認識配列を切断するとともに、認識配列のメチル化によって自己配偶子由来のmtDNAを保護する。各配偶子が持つ細胞質伝達抑制遺伝子の違いは認識配列の違いとする。
このモデルでは、認識配列に従ったDNAの切断による接合相手由来の細胞質遺伝子伝達抑制を仮定した。
ヌクレアーゼによって接合相手のミトコンドリアDNA(mtDNA)認識配列を切断するとともに、認識配列のメチル化によって自己配偶子由来のmtDNAを保護する。各配偶子が持つ細胞質伝達抑制遺伝子の違いは認識配列の違いとする。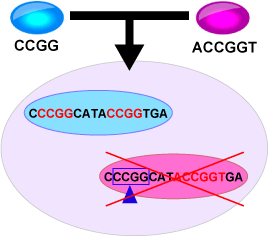 このような認識配列を持つ配偶子集団で細胞質遺伝子の片親遺伝が起こるのは図に示すような配偶子の組み合わせの時のみである。 青色で示す配偶子は"CCGG"、赤色で示す配偶子は"ACCGGT"という認識配列を持っている。これらの配偶子は認識配列に従って自己のmtDNAをメチル化している(配列中の赤い部分)。接合すると赤色配偶子由来のmtDNAは"CCGG"の部分を切断されて消滅してしまい、結果として青色配偶子由来のmtDNAが伝達される。このように一方の認識配列が他方の認識配列の一部(つまり「入れ子」構造)になっているとき片親遺伝が成立する。因に二つの配偶子が完全に同一の認識配列を持つ場合は両親遺伝し、全く異なる認識配列を持つ場合はどちらからも伝わらない。
このような認識配列を持つ配偶子集団で細胞質遺伝子の片親遺伝が起こるのは図に示すような配偶子の組み合わせの時のみである。 青色で示す配偶子は"CCGG"、赤色で示す配偶子は"ACCGGT"という認識配列を持っている。これらの配偶子は認識配列に従って自己のmtDNAをメチル化している(配列中の赤い部分)。接合すると赤色配偶子由来のmtDNAは"CCGG"の部分を切断されて消滅してしまい、結果として青色配偶子由来のmtDNAが伝達される。このように一方の認識配列が他方の認識配列の一部(つまり「入れ子」構造)になっているとき片親遺伝が成立する。因に二つの配偶子が完全に同一の認識配列を持つ場合は両親遺伝し、全く異なる認識配列を持つ場合はどちらからも伝わらない。簡単のため、認識配列をバイナリナンバーで表示しシミュレーションを行うと、以下のような直線順位が表れた。
後ろの数字(配列)が直前の数字(配列)を含んでいる。
関連文献・リンク等
リンク
- PhysarumPlus Physarum Polycephalumについて
- 東京大学大学院 植物生存システム 河野研のページ
書籍
- ミトコンドリアの謎 河野 重行著 講談社
- シリーズ進化学(3) 化学進化・細胞進化 石川 統、河野 重行、大島 泰郎、山岸 明彦著 岩波書店
- ミトコンドリアはどこからきたか 生命40億年を遡る 黒岩 常祥著 日本放送出版協会
論文
- Cytoplasmic Fusion and the Nature of Sexes
Laurence D. Hurst, William D. Hamilton
Proceedings: Biological Sciences, Vol. 247, No. 1320 (Mar. 23, 1992), pp. 189-194
back